When cells stop talking.
Greying isn't simply pigment fading out. Higher up the follicle, a reservoir of stem cells has to be told — reliably, cycle after cycle — when to stay quiet, when to wake, and when to deploy. When that local conversation weakens, the reservoir remains in place, but it can no longer be mobilised on time. The result is the slow, compounding loss of colour that most greying actually is.
Greying is a supply problem, not just a pigment problem.
Greying is described, almost always, as a loss of pigment. That description is accurate but incomplete. It names the endpoint and skips the mechanism.
The bulb — the factory at the base of each follicle — can only make colour if it is continuously restocked with fresh pigment-producing melanocytes. That restocking depends on a stem-cell reserve maintained higher up, in the bulge niche, cycle after cycle [Ref 1] [Ref 2].
What matters here is why that reserve stops behaving reliably. The answer is not simply that stem cells die. The answer is that they depend on short-range maintenance signals from neighbouring niche cells — signals that keep them alive, keep them in the right state, and trigger a controlled wave of activation at the right point in each hair cycle.
When those signals weaken or become mistimed, the reserve can still exist on paper. But it becomes harder to maintain, and harder to deploy on schedule. Fewer functional pigment cells reach the bulb. More hairs grow in with reduced pigment. The curve starts to bend, quietly, long before anything is visible in a mirror.
How cells talk.
No tissue would work if its cells could not co-ordinate. Every organ depends on cells exchanging signals that tell each other when to stay quiet, when to activate, when to divide, and when to specialise.
A signal can take several physical forms. It may be a released chemical messenger — a growth factor or a hormone. It may be a surface-bound cue that only works on direct contact between two cells. It may be a change in electrical state. It may even be mechanical — a stretch, a pressure, a shear force. The forms differ; the underlying pattern does not. A sender produces the signal, a receiver detects it through a matching receptor or sensor, and then the receiver has to decode what the input means [Ref 3] [Ref 4].
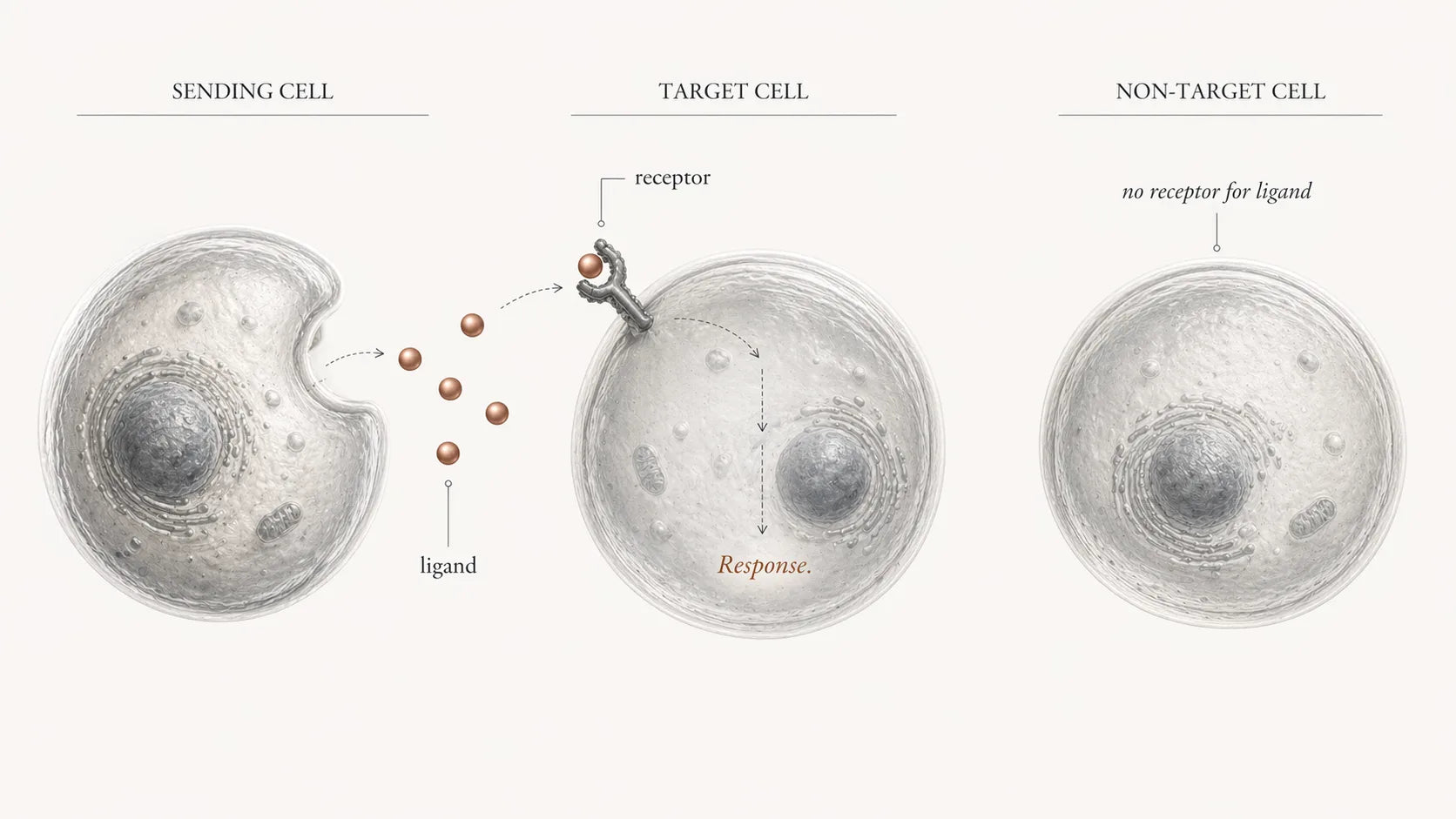
Decoding the message.
Decoding happens inside the receiving cell, through networks of molecular switches and messengers that can amplify a small external cue into a strong, specific response. The same signal can mean different things in different cell types, because the internal wiring is different. The response can also propagate outward: an activated cell may release new signals or display new surface cues that then influence its neighbours [Ref 3].
And no cell listens to only one message. Each cell integrates many signals at once, across many pathways, and its behaviour reflects the combined vote. That constant multi-signal integration is what turns simple inputs into the complex, self-stabilising communication system that keeps tissues coherent.
When that communication becomes distorted — signals too weak, too strong, mistimed, or noisy — cells start making the wrong decisions. Repair pathways can stay switched on too long. Stem cells can exit the niche on the wrong schedule. Differentiation can happen at the wrong time, or not at all.
When the greying story changed.
The turning point in greying research was the recognition that hair colour is governed upstream — at the level of the reserve — rather than downstream at the bulb [Ref 1]. That reframed the core question. It was no longer "why does the factory slow down?" It was "why does the reserve that feeds the factory stop renewing itself?"
That reframing made the niche central, because the niche is where the reserve is maintained. The niche is not a location; it is a control layer. It provides melanocyte stem cells with short-range cues for survival, for stemness, and for activation timing, so they can be deployed in a controlled wave whenever a new hair is built [Ref 2]. In practice, stem-cell maintenance and niche signalling are the same mechanism, viewed from two sides.
The conversation for colour.
Hair colour depends on a co-ordinated hand-off between two zones of the follicle. The bulb, at the base, is the production zone — where pigment is actively made during the growth phase of a hair cycle. Higher up sits the niche, where the melanocyte stem-cell reserve is maintained and prepared for the next cycle [Ref 2].
That hand-off is where signalling becomes operational. Each new hair cycle demands a fresh batch of working melanocytes. The niche has to do three things simultaneously: keep the reserve viable between cycles, hold it in the right state, and then activate a controlled fraction of it at the right time to generate pigment-cell descendants that migrate down to the bulb.
The talk is not one message; it is a layered stream of cues that the reserve integrates. Maintenance cues support survival and stress protection so stem cells don't quietly drop out. Identity cues preserve stemness and self-renewal rather than premature drift toward a wrong fate. Timing cues trigger activation and direct descendants toward the bulb when a new hair is being built.
When those cues are clear and well-timed, the reserve is stable and the bulb is reliably supplied. When they weaken or become noisy, the system still runs — it just runs less reliably. More hairs grow in with reduced pigment from one cycle to the next.
These cues are not abstract. Specific signalling pathways and molecules in the niche have been identified, and they control whether melanocyte stem cells stay maintained and when they mobilise.
The niche is a small community. Its reliability depends on both the signal sent and the signal heard. Weaken either side and the pigment pipeline becomes less reliable, cycle by cycle.
A conversation mapped: endothelin → EDNRB.
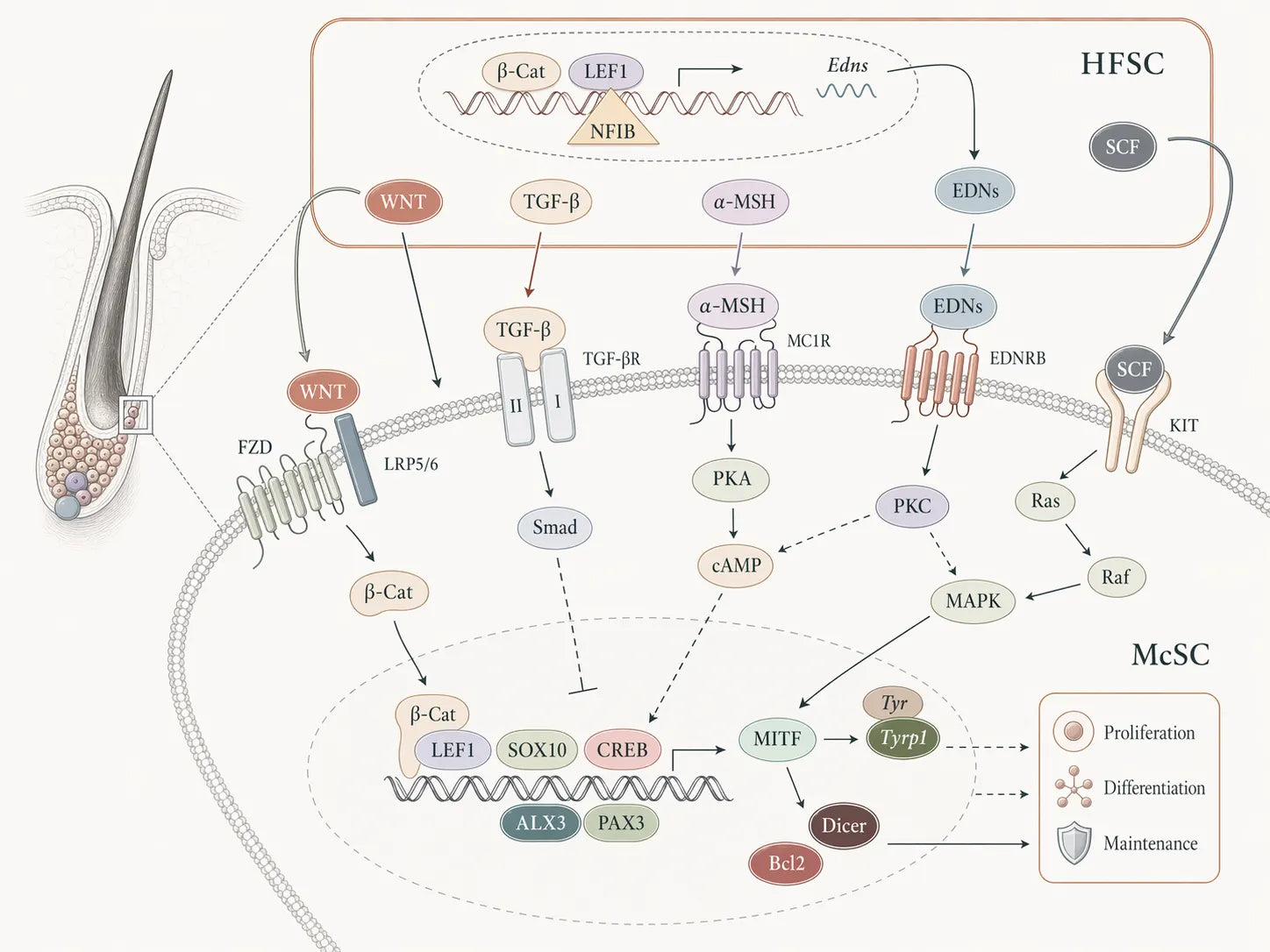
If we make niche signalling concrete, the best-studied line of dialogue is the one that runs from endothelin cues to the EDNRB receptor [Ref 8] [Ref 9].
In that configuration, niche support cells — including hair-follicle stem cells (a keratinocyte-stem-cell lineage) — sit directly next to melanocyte stem cells and behave like caretakers [Ref 2]. They release endothelins, short-range chemical messengers, into the local environment. Melanocyte stem cells in the same niche display EDNRB on their surface. When endothelin binds, the receptor fires internal pathways that support survival and stress protection, preserve stemness, and hold the reserve in a state of readiness — so a fraction can activate on time and generate pigment-cell descendants.
EDNRB signalling also connects to other niche pathways, including Wnt/β-catenin, which helps co-ordinate when melanocyte stem cells mount a regenerative response, especially after stress or injury [Ref 8].
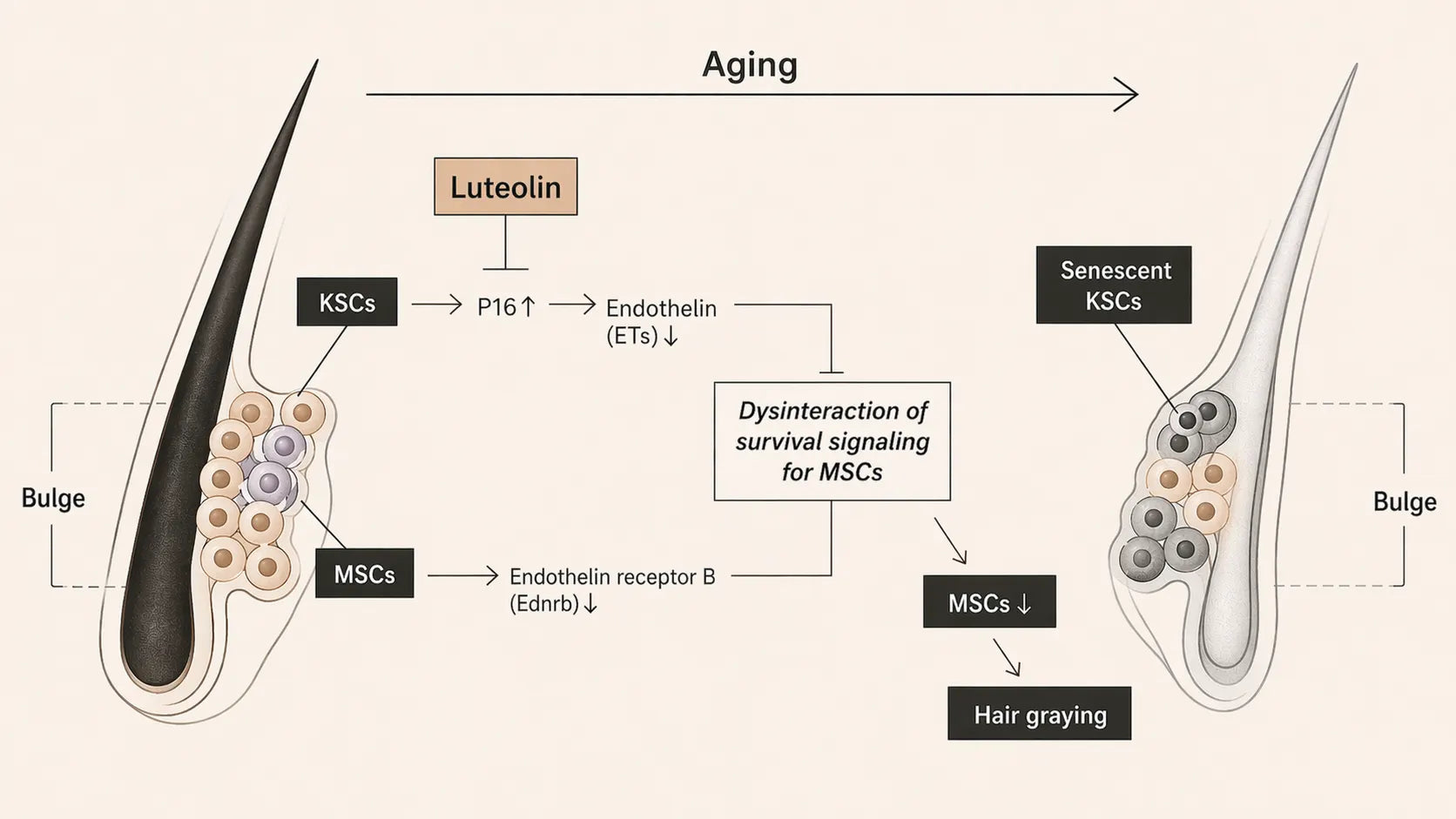
When the signal fades with age.
There is now empirical data linking changes in niche signalling to visible greying. A 2024 study in a fast-greying mouse model tested Luteolin and related flavonoids [Ref 10]; the behavioural outcomes of that work are covered in Saving the cells behind hair colour. The part that matters here is what the same study revealed about the signal itself.
In ageing mice, keratinocyte stem cells in the bulge niche showed stronger senescence markers alongside a decline in local endothelin cues. With the cue weakening, EDNRB signalling in nearby melanocyte stem cells weakened in turn — and the pigment reserve became measurably harder to maintain. In Luteolin-treated mice, those signalling measures shifted in the protective direction, consistent with a more stable niche environment and a better-preserved pigment pipeline [Ref 10].
The study also pointed to endothelin-1 as a possible mediator. In cultured human keratinocyte cells, Luteolin reduced the senescence marker p16INK4A and increased endothelin-1 gene activity. That suggests a link between less cellular senescence and more intact niche signalling. The authors flag the limitation clearly: in-vitro findings do not automatically carry over to the living human follicle [Ref 10].
Bottom line.
Signalling is what holds the pigment reserve together across decades. When the niche's local dialogue is intact, melanocyte stem cells stay alive, stay in the right state, and deploy on schedule — and the bulb gets the cells it needs to make colour. When the dialogue weakens, the reserve is still there but becomes harder to maintain and harder to mobilise. Each cycle loses a little ground. Over years, the ground loss becomes a scalp that has greyed.
Keeping hair coloured for longer is, at least in part, the same task as keeping that signal clear. The evidence so far — obtained in a model engineered to reproduce the human pathway — shows that this layer is drug-addressable: small molecules can protect the niche environment well enough to keep the conversation going, and preserving the conversation preserves the reserve. The remaining work is the human-scalp replication on a human timeline; the mechanism being replicated is already the one that matters.
- № 01 Why hair turns grey.The cascade of six compounding failures at the follicle.
- № 02 Is hair greying permanent?The half of the cascade that can still be undone.
- № 03 How stress greys hair.The nerve-driven arc that empties the pigment reserve in a single shot.
- № 04 Saving the cells behind hair colour.On Luteolin and the window to intervene before the reservoir runs out.
-
(2005). Mechanisms of hair graying: incomplete melanocyte stem cell maintenance in the niche. Science, 307(5710):720–724.Read ↗
-
(2011). Hair follicle stem cells provide a functional niche for melanocyte stem cells. Cell Stem Cell, 8(2):177–187.Read ↗
-
(2002). Cell surface receptors and their signal transduction pathways. Molecular Biology of the Cell (4th ed.), NCBI Bookshelf.Read ↗
-
(n.d.). Cell signaling. Nature Scitable.Read ↗
-
(2022). How do cells communicate? Babraham Institute.Read ↗
-
(2023). How cells talk to each other. ISTA.Read ↗
-
(2018). Regulation of melanocyte stem cell behavior by the niche microenvironment. Pigment Cell and Melanoma Research, 31(5):556–569.Read ↗
-
(2016). EdnrB governs regenerative response of melanocyte stem cells by crosstalk with Wnt signaling. Cell Reports, 15(6):1291–1302.Read ↗
-
(2017). Endothelin-3 activates melanocyte stem cells and promotes hair repigmentation after injury. Scientific Reports, 7:14284.Read ↗
-
(2024). Anti-graying effects of external and internal treatments with Luteolin on hair in model mice. Antioxidants, 13(12):1549.Read ↗